Im System der Pflanzen gehören die Leguminosae in die Abteilung der Angiospermae (bedecktsamige Pflanzen), in die Klasse der Dicotyledonae (zweikeimblättrige Pflanzen), in die Unterklasse Archiclamideae (Pflanzen, deren Kronenblätter nicht verwachsen sind), in die 23. Reihe Rosales, in die 4. Unterreihe Leguminosineae und in die Familie der Leguminosae. Diese Familie ist gekennzeichnet durch vorwiegend zygomorphe Blüten, mit meist einem Karpell, die Antheren öffnen sich fast stets mit Längsspalten, die Früchte (meist Hülsen) sind aufspringend oder geschlossen bleibend, mehr- bis vielsamig und selten einsamig.
Zur Familie der Leguminosae gehören ca. 600 Gattungen mit über 13.000 Arten, die als Bäume, Sträucher, Halbsträucher, Stauden und einjährige Kräuter aufrecht oder rankend vorkommen. Sie sind zu finden in tropischen und subtropischen Regenwäldern über Savannen und Steppen bis in Wiesen und Heiden der temperierten Gebiete und an die Grenze der Arktis. In den Tropen sind sie oft dominierende Bestandsbildner.
Die Leguminosae gliedern sich in 3 Unterfamilien, die Mimosoideae, die Caesalpinioideae und die Papilionoideae, von denen die letztere mit ihrer Schmetterlingsblüte die größte und die am meisten fortgeschrittene Gruppe darstellt. Das Gemeinsame der drei Unterfamilien ist z.B. die Ausbildung der Frucht als Hülse, was von größerer Bedeutung ist als das Trennende, so daß eine Aufteilung in drei selbständige Familien, auch weil die Grenzen nicht scharf zu erkennen sind, unberechtigt erscheint. Im ganzen stellen die Leguminosae einen sehr natürlichen Verwandtschaftskreis dar, der sich seit der Kreidezeit und dem Tertiär zu der heutigen Formenfülle entwickelt hat. Die Familie der Leguminosae ist durch die Unterfamilien Caesalpinioideae und Mimosoideae mit der Familie der Chrysobalanaceae und durch andere Artengruppen mit den in den Tropen weitverbreiteten Conneraceae verwandt. Aber die Ausbildung der Frucht ist, wie gegen die Familie der Krameriaceae, ein sicheres Unterscheidungsmerkmal.
Alle landwirtschaftlich wichtigen Arten, die verschiedenen
Tribus und Gattungen angehören und für die Körnernutzung
Bedeutung erlangt haben, sind in der Unterfamilie der Papilionoideae
zu finden.
Unterfamilie Papilionoideae (Schmetterlingsblütler)
Tribus: Genisteae (Ginsterartige)
Wurzel
Die Wurzel der Leguminosae ist meist eine kräftige,
tief in den Boden eindringende Pfahlwurzel mit zahlreichen, mehr oder weniger
verzweigten Seitenwurzeln. FRUWIRTH unterscheidet drei Wurzeltypen:

Darstellung 1: Wurzeltypen von Leguminosen (nach FRUWIRTH))
Typ 1 bildet eine lange, wenig verzweigte Hauptwurzel, die tief in den Boden eindringt und Wasser und Nährstoffe aus den unteren Schichten nutzen und hervorholen kann = Lupinus-Typ.
Typ 2 hat eine weniger tiefgehende Pfahlwurzel, dafür aber stärkere und weiter verbreitete Seitenwurzeln = Pisum-Typ.
Typ 3 umfaßt Pflanzen, die zwar auch eine Hauptwurzel ausbilden, daneben aber kräftige Seitenwurzeln, die sich wiederum mehrfach verzweigen = Phaseolus-Typ.
Auch kommen bei einigen Arten Wurzelknollen vor, die als Nährmittel genutzt werden können.
Die Leguminosenwurzeln haben ein bedeutendes Aufschließungs- und Aufnahmevermögen für Bodennährstoffe und Wasser. Sie können deshalb Kulturbringer für schwere und steinige Böden, aber auch für leichte Sandböden sein. Ihre Stoppelrückstände reichern die Krume mit Nährstoffen aus tieferen Schichten an oder ermöglichen überhaupt einen Kulturpflanzenbau. Die Leguminosen sind bereits hierdurch wertvolle Vorfrüchte für andere Kulturpflanzen.
Die größte Bedeutung haben jedoch Leguminosenwurzeln
durch die Symbiose mit Bakterien (Bacterium radiuzola
Prazuiquski [= Rhizobium leguminosarum Frank.]), die in die
Wurzeln eindringen und artspezifische Knöllchen an den Haupt- und
(oder) Nebenwurzeln ausbilden, die Knöllchenbakterien (s. Abschnitt
II). Die Bakterien entnehmen der Pflanze mineralische Nährstoffe,
die Pflanze nutzt dafür die stickstoffreichen Ausscheidungen der Bakterien,
da letztere Stickstoff aus der Luft entnehmen können. So sind die
Hülsenfruchtarten fähig mit Hilfe ihrer Knöllchenbakterien
Stickstoff aus der Luft für sich und nachfolgende oder daneben wachsende
Pflanzen zu nutzen (HILTNER). Die symbiotische Beziehung zwischen Bakterien
und Wirtspflanze ist wechselseitig artspezifisch, wie folgende Zusammenstellung
zeigt:
Rhizobium-Gruppen wichtiger Leguminosae (nach
REHM & ESPIG)
| Rhizobium-Art | Leguminosen-Gattungen |
| Rhizobium meliloti | Medicago, Melilotus |
| Rhizobium trifolii | Trifolium |
| Rhizobium leguminosarum | Lathyrus, Lens, Pisum, Vicia |
| Rhizobium phaseoli | Phaseolus |
| Rhizobium lupini | Lupinus |
| Rhizobium japonicum | Glycine |
| Rhizobium sp. | Cicer |
| Rhizobium sp. | Alysicarpus, Arachis, Cajanus, Canavalia, Crotalaria, Cyamopsis, Desmodium, Indigofera, Lespedeza, Mucuna, Pueraria, Vigna |
| Rhizobium sp. | Lotononis |
| Rhizobium sp. | Lotus |
Sproß
Die Stengel der Leguminosen sind oft vierkantig
oder rund mit mehr oder weniger deutlichen Riefen oder geflügelt.
Für die Wuchsform unterscheidet HEGI folgende Achsenformen:
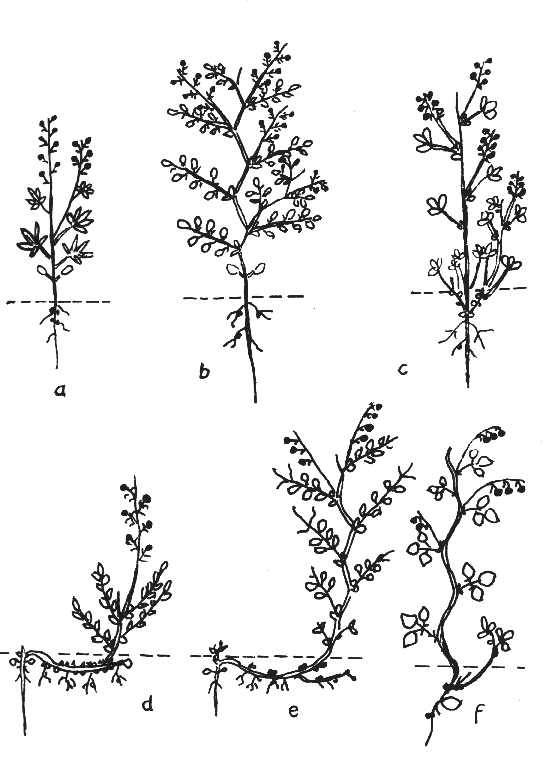
Darstellung 2: Schematische Darstellung des Sprossaufbaues krautiger Papilionoideae
Die Achsen erster und dritter Ordnung sind schwarz, diejenigen zweiter und vierter Ordnung hell gehalten. a: Zweiachsige Pflanze (Lupinus). b: Dreiachsige Pflanze mit Langsproß und Stipellen (z.B. Galegeaen). c: Dreiachsige Pflanze mit Erdstock (z.B. viele Trifolieaen). d: Vierachsige Pflanze mit Wurzelstock (z.B. Hedysarum). e: Vierachsige Pflanze mit Bodenausläufern und Blattranken (Vicieaen). f: Vierachsige Pflanze mit windendem Stengel (Phaseoleaen) (nach HEGI).
Die Laubblätter können unpaarig, dreizählig, gefingert sein, oder ein- bis vielpaarige Fiederblättchen tragen. Die Blätter und Blättchen sind verschieden gestaltet. Die endständigen Fiederblättchen sind entweder als Blättchen oder verändert als Ranken, auch als Spitzen ausgebildet.
Die Form der Blättchen ist meist oval, lanzettlich, verkehrt eiförmig mit glatten, gewellten oder gezähnten Rand. Sie stehen paarig oder unpaarig an einem runden oder kantigem Blattstiel.
Die Nebenblätter (Stipulae, Stipeln,
Stipellen) sind Ausgliederungen des Balttgrundes. Durch ihre Verschiedenheit
in Größe, Form, Farbe und Behaarung haben sie Bedeutung als
wichtiges Merkmal und für die Funktion. Je nach ihrer Größe
sind sie mehr oder weniger an der Assimilation beteiligt und können
in extremen Fällen, z.B. bei Lathyrus aphaca, diese fast ganz
übernehmen, da die Blättchen zu Ranken mit verminderter assimilationsfähiger
Fläche umgebildet sind. Ein anderes Beispiel für die Vielgestaltigkeit
sind die Dornen bei Robinia pseudoacacia.
Bestimmungsschlüssel für die Blätter
wichtiger Leguminosae (nach STÄHLIN):
Gruppe I: Blätter ungeteilt
Canavalia gladiata (Jacq.) DC.
Cyamopsis tetragonolobus (L.) Taub.
Glycine soja (L.) Sieb. et Zucc.
Macrotyloma geocarpum (Harms) Maréchal et Baudet
Macrotyloma uniflorum (Lam.) Verdc.
Pachyrrhizus erosus (L.) Urban
Psophocarpus tetragonolobus (L.) DC.
Lotus tetragonolobus Moench = Tetragonolobus purpureus L.
Sphenostylis stenocarpa (Hochst. ex A. Rich.) Harms
Vigna aconitifolia (Jacq.) Maréchal
Vigna angularis (Willd.) Ohwi et Ohashi
Vigna umbellata (Thunb.) Ohwi et Ohashi
Vigna unguiculata (L.) Walp.
Die Blüten stehen einzeln, zu zweit oder in traubigen Blütenständen. Der Blütenbau ist sehr unterschiedlich und nicht einmal innerhalb der Gattung konstant (HEGI). Der Kelch ist zweispaltig oder meist glockig und mehr oder weniger deutlich fünfzähnig.
Die Blüten der Leguminosae und speziell der Papilionoideae (Schmetterlingsblütler) sind zygomorph, d.h. zweiseitig symmetrisch, wobei deren Symmetrieachse zwei spiegelbildlich gleiche Hälften besitzt. Selten kommen aktinomorphe, d.h. vollkommen strahlige mit radiär regelmäßigem Blütenbau vor. Die Blüte besteht aus fünf Kronenblättern (Petalen). Bei der zygomorphen Blüte sind die beiden unteren Kronenblätter teilweise zu einem Schiffchen (Carina) verwachsen; dies ist oben ganz oder nur an der Spitze offen. Im Schiffchen liegt der Griffel von den 10 Staubgefäßen umhüllt, die, alle 10 oder nur 9, eine Röhre mit Honigdrüsen bilden. Zwei weitere Blütenblätter bilden die Flügel (Alae), die mit dem Schiffchen durch Wülste, Leisten, Zähne oder Überwölbungen so verbunden sind, daß das Schiffchen mit ihnen bewegt wird. Die Fahne (Vexillum) ist fast immer groß ausgebildet und lebhaft gefärbt. Durch unterschiedliche und intensive Färbung der verschiedenen Blütenteile entsteht eine starke Anlockung der Insekten, aber auch vielfach durch Duftstoffe, die vom Honig im Schiffchen ausgehen.
Durch die frühe Reife von einem Teil der Staubgefäße
herrscht Selbstbefruchtung schon in der geschlossenen Knospe vor. Einige
Arten sind jedoch auch überwiegende Fremdbefruchter durch Insekten,
wobei der besondere Bau des Schiffchens mit der "Geschlechtssäule"
die nektarsammelnden Hummeln und Bienen reichlich mit Pollen einstäubt
(s. Darst. 3).
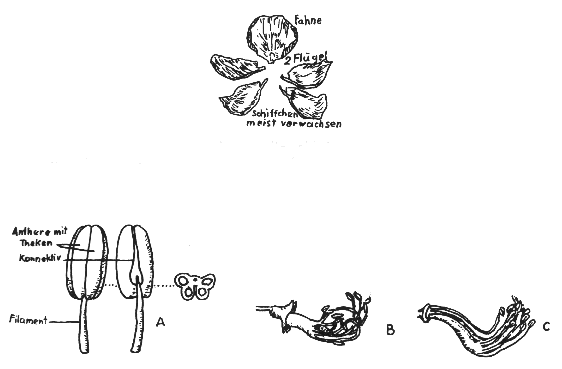
Darstellung 3: Blüten nach Entfernung der Petalen (Kronblätter) (nach ENGLER)
B: Filamente verwachsen mit geschlossener Filamentröhre, Stamina (10)
C: Filamente verwachsen mit nach hinten offener Filamentröhre, Stamina (9) + 1 Vexillastamen frei
1. Schmetterlingsblüten mit Klappvorrichtung: z.B. Trifolium pratense.
2. Blüten mit unterschiedlicher Reife der Antheren innerhalb der Blüte und dazu mit Pumpvorrichtung: z.B. Lupinus, Lotus tetragonolobus.
3. Schmetterlingsblüten mit Explosionsvorrichtung: z.B. Medicago sativus.
4. Blüten mit Bürstenvorrichtung: z.B. Pisum sativum.
Nach der Befruchtung sterben die Kronenblätter schnell ab, der Fruchtknoten beginnt zu wachsen und bildet die Hülse mit den Samen.
Frucht
Die Früchte sind als Hülsen ausgebildet und aus einem Karpell entstanden. Bei den zweiklappigen Hülsen ist die Entwicklung besonders gut zu erkennen. Sie haben eine dorsale und eine ventrale Naht. Die beiden Ränder der ventralen Naht sind deutlich verstärkt und haben sich aus den Plazenten entwickelt. Die Samenanlagen stehen sich an den beiden Rändern alternierend gegenüber und sind mit der Plazenta durch einen kurzen Stiel, dem Funikulus, verbunden. Die dorsale Naht ist bereits beim postfloralen Karpell vorgebildet worden. Der eigentliche Hülsentyp springt zur Reife an beiden Nähten auf, was eine Besonderheit der Leguminosae ist. Damit sind sie von den anderen Familien mit Springfrüchten zu unterscheiden. Die Hülsen lassen sich in zwei Typen unterteilen:
I. Der eigentliche Hülsentyp (Springfrucht) mit mehreren Samen ist längsgeteilt und springt bei der Reife an beiden Nähten zweiklappig bis zum Stielansatz auf.
II. Der geschlossen bleibende Typus teilt sich auf in die ein- oder wenigsamige Schließfrucht (Balg- oder Schließfrucht), bei Reife nicht aufspringend oder mit einem Deckel sich öffnend und in die vielsamige, durch Querwände abgeteilte Gliederhülse, deren Glieder je einen Samen enthalten, nach der Reife auseinanderbrechend, ohne Samen freizugeben.
Diese Typen kommen bei den Tribus wie folgt vor:
Genistae: 2-klappige Hülsen (bei Arachis
hypogaea nicht aufspringend,
da in der Erde)
Gliederhülsen
Onoideae: 2-klappige Hülsen
Phaseoleae: meist 2-klappige Hülsen
Robineae: 2-klappige Hülsen
Trifolieae: Hülse 1- bis 2-samig, kugelig oder
mehr oval mit Spitze
Hülse 1-samig mit Deckel
Hülse 1- bis vielsamig von verschiedener Gestalt
(kugelig, oval mit Spitze, diskusartig, sichelförmig, einfach bis
mehrfach gewunden) leicht aufspringend
Vicieae: 2-klappige Hülsen
Samen
Aus einer Samenanlage (Ovulum) im Fruchtknoten (Ovarium) entwickelt sich der Samen. Man erkennt am Samen die Mikropyle (Keimmund = Kanal, durch den die Pollenkerne auf die Spitze des Nucellus gelangen), durch die bei der Keimung das Keimwürzelchen und das Hypokotyl mit den Kotyledonen (Keimblätter) austreten. Ferner ist die Abbruchstelle des Funikulus (Nabelstrang) zu sehen, die als Hilum (Nabel) besonders erkennbar wird. Bei anatrophen bis kampylotrophen Samenanlagen der Leguminosae ist am Samen ein Streifen erkennbar, der vom Hilum ausgeht und als Raphe (Samennaht) bezeichnet wird. In der Raphe verläuft das vom Funikulus ausgehende Leitbündel. Vielfach ist eine punktförmige oder dreieckige, oft dunkler gefärbte Erhebung, das Strophiolum (Samenschwiele = Ganderschwiele), ausgebildet. Außerdem kommen Arillusbildungen (Arillus = Samenmantel) vor (s. Darst. 4).
Im allgemeinen (außer bei Orchideae) enthält
der Samen Nährgewebe. Die Speicherung der Reservestoffe erfolgt
im Embryosackgewebe, das in dieser Funktion kurz als Endosperm bezeichnet
wird, oder im Nucellus. Der Embryo bleibt bei diesen Samentypen klein.
Anders ist es u.a bei den Leguminosae, die das gesamte Nährgewebe
kaum bis garnicht besitzen, sondern die Reservestoffe vor allem in den
Kotyledonen (Keimblättern) speichern.
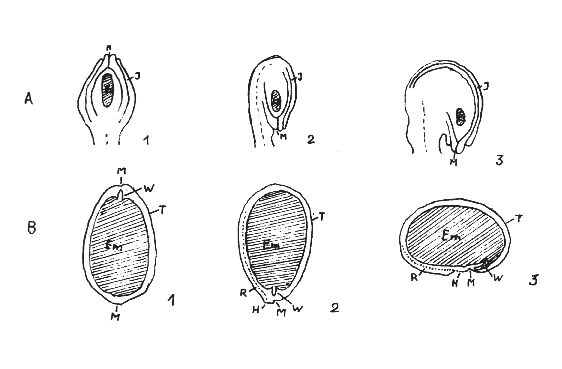
Darstellung 4: Samenanlagen und Samenformen (nach ENGLER und TROLL)
B: Samenformen M= Mikropyle, H= Hilum (Nabel), R= Raphe, T= Testa aus, J= Integumente entstanden, W= Radicula (Keimwurzel), Es= Embryosack bei Samenanlagen schraffiert, Em= Embryo schraffiert.
Die Testa (Samenschale) bildet sich aus den Integumenten der Samenanlage. Sie ist lederartig bis fast knöchern mit charakteristischer Palisaden- und Trägerschicht (selten dünn und fleischig, was bei den Mimosoideae vorkommt), meist glatt, matt oder glänzend, zuweilen auch körnig oder warzig und häufig lebhaft pigmentiert, einfarbig oder mehrfarbig gefleckt, marmoriert oder punktiert.
Der Nabel (Hilum) ist scharf umgrenzt und meist vom Funikulus (Nabelstrang) glatt abgebrochen. Seine Größe und Form ist verschieden und wird als Unterscheidungsmerkmal der Gattungen herangezogen. Bei den Papilioideae ist er relativ groß mit einer Längsfurche, kreisförmig bei Lotus und Trifolium, oval bei Cicer und Pisum, länglich-oval bei Phaseolus, lineal bei Dolichos, strichförmig bei Vicia, vertieft bei Lupinus. Gelegentlich wird der Nabel durch eine anders gefärbte Zone der Testa oder einem Wulst hervorgehoben.
An einem Ende des Nabels ist die Mikropyle inseriert,
die oft einen deutlichen Vorsprung durch die Spitze des Würzelchens
zeigt; am anderen Ende ist die Raphe (Samennaht) erkennbar (s. Darst. 5).
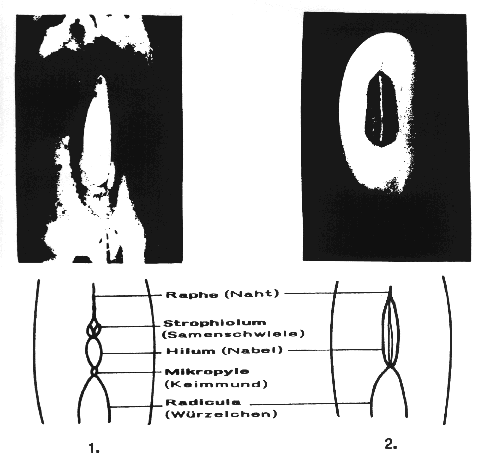
Darstellung 5: Schematische Nabelpartien von 1. Phaseolus vulgaris und 2. Glycine max (nach ZADE)
Der Verbreitung der Samen dient u.a. der Arillus (Samenmantel), der die Samenschale mehr oder weniger umgibt und verschiedener Herkunft ist. Arillusbildungen fehlen bei den Trifolieae- und Loteae- Arten und sind bei den Vicieae- und bei Phaseolus sp. stets vorhanden.
Das gerade, meist schiefe oder eingekrümmte
Würzelchen liegt bei den Keimblättern mehr oder weniger dicht
an, es ragt gewöhnlich etwas vor und ist dann mitbestimmend für
die Form des Samens (s. Darst. 6).

Darstellung 6: Samen einiger Körnerleguminosen (nach FRUWIRTH)
1. a. Vicia faba major, Große Ackerbohne; 1. b. Vicia faba minor, Kleine Ackerbohne; 2. Vicia narbonensis, Schwarze Ackerbohne; 3. Vicia sativa, Saatwicke; 4. Vicia ervilia, Ervilie; 5. Vicia articulata, Wicklinse; 6. a. Phaseolus vulgaris, Gartenbohne; 6. b. Phaseolus vulgaris, rundovale Gartenbohne; 7. Phaseolus coccineus, Feuerbohne; 8. a. Phaseolus lunatus var, macrocarpus, Limabohne; 8. b. Phaseolus lunatus var. lunatus, Mondbohne; 9. Vigna unguiculata ssp. unguiculata, Augenbohne; 10. Vigna unguiculata ssp. cylindrica, Angola-Erbse; 11. Vigna unguiculata ssp. sesquipedalis, Spargelbohne; 12. Vigna radiata, Mungbohne; 13. Lupinus albus, Weiße Lupine; 14. Lupinus luteus, Gelbe Lupine; 15. Lupinus angustifolius, Blaue Lupine; 16. Lablab purpureus, Helmbohne; 17. Pisum sativum, Erbse; 18. Lathyrus sativum, Eßbare Platterbse; 19. Cicer arietinum, Kichererbse; 20. Lens esculentum, a. Hellerlinse, b. mittlere Linse; 21. Glycine max, Sojabohne; 22. Lotus tetragonolobus, Rote Spargelerbse.Die Reife der Samen erfolgt meist über der Erde, bei einigen Arten jedoch in der Erde (z.B. Arachis hypogaea, Vigna subterranea).
Die Keimung verläuft hypogäisch (Keimblätter bleiben in der Erde) oder epigäisch (kommen mit dem Sproß aus der Erde).
Äußere Merkmale der
zur Kornnutzung geeigneten Gattungen von Hülsenfrüchtlern
(nach HEGI, ENGLER, ENGLER & PRANTL, MANSFELD
sowie RACHIE & ROBERTS)
Wie bereits ausgeführt, gehören die in diesem Buch besprochenen Arten alle zu der Unterfamilie der Papilionaceae und innerhalb dieser zu denen, die eine mehr oder weniger deutliche zygomorphe, schmetterlingsförmige Blüte haben. Die 10 Stamina (Staubblätter) sind mehr oder weniger stark verwachsen und sind entweder zu (9) gebündelt + 1, zu (10) gebündelt oder zu (5) + (5) jeweils gebündelt. Die Arten lassen sich nach ENGLER wie folgt einordnen:
Tribus Genisteae: Stamina (10), selten (9) + 1 oder (5) + (5).
Gattung Lupinus L. mit ca. 200 Arten.
Meist kräftige, zweiachsige Kräuter mit aufrechten Stengeln, selten strauchig; Wurzeln verzweigt, tiefgehend mit großen Knöllchen reichlich besetzt; die langgestielten Laubblätter fast immer fingerförmig mit 3 bis 15 ganzrandigen Blättchen; die spitzen Nebenblätter sind zur Blattscheide verwachsen; die großen Blüten stehen in endständigen, aufrechten Trauben oder Ähren; die Krone ist sehr unterschiedlich gefärbt; die Fahne ist kreisrund bis oval; die Flügel am oberen Rand faltig bis runzelig; das geschnäbelte Schiffchen ist gekrümmt; die Antheren sind sehr ungleich und kahl; der Fruchtknoten ist sitzend mit mehreren Samenanlagen; die zweiklappigen Hülsen sind meist groß, ledrig, zusammengedrückt und oft stark behaart; die Samen variieren stark in Größe und Färbung, sie sind rundlich meist etwas abgeflacht; der ovale Nabel ist ohne Nabelwulst.
Meist ausdauernde, selten einjährige Kräuter und Halbsträucher; die ganzrandigen Laubblätter sind drei- bis fünfzählig, unpaarig gefiedert, die Spindel in einem Blättchen, niemals in einer Ranke oder Granne endend; meist sind 9 Staubblätter verwachsen, selten alle 10; die mittelgroßen Blüten stehen in den Blattachseln, sie sind zwei- oder einblütig bis doldenförmig, mit Nudelpumpeinrichtung, gelb oder rot gefärbt; der sitzende Fruchtknoten enthält viele Samenanlagen; die Hülsen sind vielförmig, gebogen oder gerade, stielrund, kantig oder vierflügelig; die Samen sind rund und mehr oder weniger abgeflacht.
Gattung Arachis L. mit 12 Arten.
Primitiv- und Kulturpflanzen mit liegenden, halbaufrechten und aufrecht wachsenden Stengeln, Blütenstände mit einer oder nur wenigen Blüten; Blüten mittelgroß, leuchtend gelb; nach der Befruchtung bildet sich ein in den Boden wachsender Fruchtstiel (Karpophor), an dessen Spitze sich im Boden die Frucht entwickelt und ausreift; die Frucht ist eine derbe, stark genetzte Hülse mit 1 bis 4 rot, braun oder violett gefärbten Samen mit hohem Eiweiß- und Ölgehalt.
Einjährige, selten mehrjährige kleine bis mittelgroße Kräuter mit starkem, aromatischem Geruch; Laubblätter dreizählig mit meist gezähnten Blättchen, nie mit Stipellen; das mittlere Blättchen deutlich gestielt; Nebenblätter meist häutig, oft mit dem Blattstiel verbunden; die Blüten sind meist klein, in Köpfchen oder kurzen Trauben in den Blattachseln stehend; die Kronblätter sind blaßgelb bis dunkelgelb oder blau gefärbt; das Schiffchen nicht eingerollt; der obere Staubfaden nicht verwachsen; der Fruchtknoten ist sitzend oder kurz gestielt; die Hülsen sind meist hornförmig, gerade oder schwach gekrümmt; die Samen sind sehr unterschiedlich in Größe, Form und Oberflächenausbildung.
Einjähriges Kraut mit gezähnten, stark stieldrüsigen Blättchen. Laubblätter mehrjochig gefiedert oder gefingert, die Spindel in einem Blättchen endend, die unteren Blättchen nicht nebenblattartig; Staubblätter meist 9, selten alle 10 verwachsen; die Blüten stehen einzeln in den Blattachseln.
Gattung Vicia L. mit 150 - 200 Arten.
Einjährige und ausdauernde Kräuter mit meist schlaffen, kletternden oder selten aufrechten Sprossen; oft mit Bodenausläufern ohne Rhizome oder Bodenknollen; Laubblätter überwiegend zweizeilig, paarig gefiedert, meist mehrjochig, selten nur ein- oder zweijochig mit einer verzweigten Wickelranke, selten Spindel in einer kurzen Granne endend; Nebenblätter meist halbpfeilförmig, ganzrandig oder gezähnt, häufig auf der Unterseite extra-florale Nektarien; Blütenstände stets aus Blattachseln wachsend, vielförmig, immer in einseitigen Trauben oder in ein- oder dreiblütigen, kurz gestielten Büscheln stehend; Blüten bis etwas über 3 cm lang, abstehend oder nickend, meist an kurzen Stielen stehend; Krone mit einem kurzen bis ziemlich langen Nagel, blauviolett, purpur, weißlich oder hellgelb gefärbt; Hülsen häutig bis lederartig, abgeflacht bis fast rund, meist bei Reife aufspringend; Samen in Form, Farbe und Oberfläche sehr verschieden.
Gattung Pisum L. mit 6 - 7 Arten.
Pflanze völlig kahl, ein- oder zweijährig; Nebenblätter größer als die Blättchen, eiförmig bis herzförmig; Laubblätter teilweise in Ranken umgewandelt; die 1- bis 3blütigen Blütenstände so lang wie oder kürzer als die Nebenblätter; Blüten 2 bis 3 cm lang, weiß oder bunt, rot, rosa, violett; Staubfadenröhre gerade abgeschnitten; Griffel flach, zusammengefaltet; Hülsen zwei- bis vielsamig; Samen vielförmig: rund, glatt, runzelig, eingedellt, klein = 0,2 bis 0,3 cm bis groß = 0,8 bis 1,2 cm, grün, gelb, braun teilweise mit dunkler Zeichnung.
Gattung Lens Miller mit 6 Arten.
Ein- oder zweijähriges Kraut mit scharfkantigen Stengeln und schmalen elliptischen Blättchen; Laubblätter paarig gefiedert, Blattspindel oft in einer Ranke endend; Staubblätter meist zu einer Röhre verwachsen, eines freistehend; Griffel im oberen Teil meist behaart; Blüten klein bis mittelgroß, weißlich bis bläulich, in langgestielten ein- bis dreiblütigen Trauben stehend; Hülsen kurz, flach, in Größe und Farbe stark variierend.
Gattung Lathyrus L. mit 160 Arten.
Kräuter; Stengel häufig geflügelt; Laubblätter mehrjochig gefiedert oder gefingert, selten auf Ranke oder Nebenblätter reduziert oder nur einjochig gefiedert, Blattspindel oft in einer Ranke endend; Staubblätter meist 9, selten alle 10 verwachsen; Griffel im oberen Teil meist behaart, oft abgeflacht oder gedreht; Nebenblätter kleiner als die Blättchen, meist halbspießförmig, nie mit Drüsenfleck und nie stärker gezähnt; Blüten in langgestielten, meist wenigblütigen Trauben, sehr verschieden gefärbt.
Aufrechte Kräuter mit unpaarig gefiederten Blättern, die aus 3 bis 7 Blättchen bestehen. Die Nebenblätter sind borstenförmig, klein. Die kleinen, achselständigen Blüten stehen in lockeren Trauben. Der breite, schiefstehende Kelch hat unten längere Zähne; die Fahne ist verkehrt-eiförmig, sitzend. Die freistehenden Flügel sind länglich; das Schiffchen ist leicht aufwärts gebogen und stumpf. Die Staubblätter sind in eine geschlossene Röhre verwachsen. Der sitzende Fruchtknoten mit 6 bis 10 Samenanlagen trägt einen kurzen, oben etwas gekrümmten Griffel mit kopfförmiger Narbe. Die etwas vierkantigen Hülsen sind spitz zulaufend und innen etwas gefächert.
3 Arten werden kultiviert: C. tetragonoloba (L.) Taub. mit aus 3 gezähnten Blättchen bestehenden Blättern; in Afghanistan bis Vorderindien verbreitet. C. senegalensis Guill et Perr. mit aus 3 bis 7 ganzrandigen Blättchen bestehenden Blättern; im nördlichen tropischen Afrika und in Arabien angebaut. C. serrata Schinz im Amboland (im nördlichen Südwestafrika) verbreitet.
Gattung Glycine L. mit ca. 60 Arten.
Fahne und Schiffchen wenig unterschiedlich in der Länge, Fahne nach oben gerichtet; Hülsen über dem Boden reifend. Blütenstände meist Büschel von wenigen kleinen Blüten oder sehr kurze Trauben, meist geschlossen abblühend; Griffel im oberen Teil kahl; Wild- und Nutzpflanzen mit schwach windendem, als Kulturpflanzen aufrechtem Stengel; Blüten mit weißen bis lila gefärbten Vorblättern; oberstes Staubblatt in der Mitte mit den anderen verwachsen; Hülsen etwas gedunsen; Samen 4 bis 10 mm lang, meist einfarbig mit linealem Nabel ohne Nabelwulst.
Subtribus Galactiinae: Blütenstandsachse an der Ansatzstelle der Blüten knotig verdickt; Griffel kahl; Petalen fast gleich lang.
Gattung Canavalia DC. mit ca. 50 Arten.
Windende oder niederliegende Kräuter mit gefiederten Blättern mit je 3 Blättchen und Stipellen. Die Nebenblätter sind klein, manchmal auch warzen- oder drüsenförmig. Die meist großen, weißlich, rosenrot oder violett-purpurn gefärbten Blüten stehen achselständig in Trauben, einzeln, paarweise oder in Büscheln. Der Kelch ist zweilippig mit großem Ober- und kleinem Unterteil. Die große, runde bis eiförmige Fahne ist zurückgebogen und trägt am Grunde oft Öhrchen. Die Flügel sind schmal, sichelförmig oder etwas gedreht. Das Schiffchen ist breiter als die Flügel, stumpf, teilweise mit nach innen gedrehtem Schnabel. Der Fruchtknoten ist sehr kurz gestielt oder sitzend. Der Griffel ist eingebogen oder mit dem Schiffchen eingerollt mit kleiner Narbe. Die Hülsen sind länglich bis breit zusammengedrückt, oft etwas aufgedunsen, die obere Naht ist oft doppelt geflügelt oder nur als Längsrippe vorstehend. Die Samen sind groß, eiförmig bis rund, etwas abgeflacht.
Es sind 12 Arten in den wärmeren Gebieten beider Erdhälften bekannt. Vor allem sind Canavalia ensiformis DC., Canavalia gladiata DC., Canavalia obtusifolia DC. und Canavalia bonariensis Lindl. verbreitet.
Subtribus Cajaninae: Blütenstandsachse an der Ansatzstelle der Blüten nicht knotig verdickt; Blättchen meist ohne Stipellen; Pflanzen besonders die Blattunterseite mit Drüsenpunkten, Griffel kahl.
Gattung Cajanus DC. mit 11 Arten.
Aufrechter Halbstrauch mit gefiederten Blättern und je 3 Blättchen; lange pfriemförmige, frühzeitig abfallende Nebenblätter. Die gelben, meist purpurn gestreiften Blüten stehen achselständig in gestielten Trauben. Der Kelch ist glockig mit spitzen Zipfeln. Die runde Fahne ist zurückgebogen und trägt am Grunde nach innen gebogene Öhrchen; die schiefstehenden Flügel sind verkehrt-eiförmig; das Schiffchen hat eine stumpfe, eingebogene Spitze. Die Hülsen sind schief zugespitzt und zusammengedrückt mit Querlinien. Die fast kugeligen Samen sind leicht eingedellt und ohne Nabelwulst. - In den Tropen beider Hemisphären angebaute und verwilderte Arten. Einheimisch wahrscheinlich in Afrika.
Subtribus Phaseolinae: Blütenstandsachse an der Ansatzstelle der Blüten knotig verdickt; Griffel längs der Innenseite gebärtet oder um die Narbe herum pinselförmig behaart; selten kahl.
Gattung Phaseolus L. mit ca. 200 Arten.
Schlingpflanze oder einjährige, aufrechte Kräuter. Stengel höchstens am Grunde verholzt; Sprosse mit Kletterhaaren; Laubblätter mit langen Stielen, drei großen netznervigen Blättchen und vier kleinen Stipellen am Grunde der Blättchen; Nebenblätter klein, meist ganzrandig; die Blüten stehen aufrecht oder abstehend in Trauben in den Blattachseln mit je zwei, meist großen Vorblättern unter jeder Blüte; der Kelch ist glockig oder hat eine kurze Röhre; die stets asymmetrische Krone ist weiß, rot, violett oder gelb gefärbt; die Fahne kreisrund, oft gewellt oder verbogen, nur wenig länger als die verkehrt eiförmigen Flügel; das Schiffchen hat einen stumpfen, spiralig eingerollten Schnabel; das oberste Staubblatt ist frei; der Fruchtknoten ist fast sitzend und mit einem becherförmigen Diskus umgeben, er enthält viele Samenanlagen; Hülsen und Samen sehr vielförmig.
Gattung Vigna Savi mit ca. 150 Arten.
Der Gattung Phaseolus steht Vigna Savi sehr nahe; bis zum Bekanntwerden der amerikanischen Phaseolus-Arten wurden die Vigna-Arten allgemein als Phaseolus bezeichnet; Vigna unterscheidet sich von Phaseolus hauptsächlich durch die symmetrischen Blüten.
Wegen der Nutzung von Hülsen und Samen angebaute Arten; Hülsen reifen über dem Boden; die Blüten stehen meist in Trauben mit knotigen Verdickungen am Ansatz der Blütenstiele; das Schiffchen ist nicht eingerollt; der Griffel ist im oberen Teil völlig kahl und das oberste Staubblatt steht ganz frei; Samen meist unter 1 cm groß; Nabel keilförmig, eingesenkt oder vortretend, oft schwarz umrandet; Nabelwulst teilweise schwach entwickelt, oft undeutlich.
Gattung Pachyr(r)hizus Rich. ex DC. mit 6 Arten.
Hohe, windende Kräuter; dreifach gefiederte Blätter mit Stipellen und oft gebuchteten oder gezähnten Blättchen. Die Blüten stehen in achselständigen, verlängerten bis rispigen Trauben. Bei dem glockenförmigen Kelch sind die oberen beiden Zipfel verwachsen. Die Fahne ist breit, verkehrt-eiförmig mit nach innen gebogenen Öhrchen. Die länglichen, gebogenen Flügel sind ebenso lang wie das stumpfe, eingebogene Schiffchen. Der Fruchtknoten mit sehr kurzem Stielchen hat viele Samenanlagen. Der an der Innenseite behaarte Griffel ist verdickt und hat eine breite, etwas eingerollte Spitze und eine fast kugelige Narbe. Die flachen Hülsen mit Querleisten sind linear und die Samen eiförmig.
Drei Arten werden kultiviert: P. erosus (L.) Urban (= P. bulbosus (L.) Britton) im tropischen Amerika und in Asien häufig angebaut; P. tuberosus (Lam.) Spreng. ist wahrscheinlich im Gebiet des oberen Amazonas beheimatet. P. palmatilobus (Moç et Sessé) Benth. ist in Mexiko verbreitet. Die großen, knolligen Wurzeln werden zu Speisen verarbeitet.
Gattung Dolichos L. mit ca. 120 Arten.
Windende, niederliegende oder aufrechte Kräuter oder Halbsträucher mit gefiederten, drei Blättchen tragenden Blättern mit Stipellen, Nebenblätter meist klein. Die Blüten stehen einzeln, gebüschelt oder in Trauben in den Blattachseln; die Blütenfarbe ist gelblich bis weißlich, fleischfarben oder violett. Der Kelch ist glockenförmig. Die runde Fahne trägt am verdickten Grund nach innen stehende Öhrchen. Die eiförmigen Flügel sind gekrümmt und hängen dem oft eingebogenen Schiffchen an. Der Griffel ist verdickt, meist etwas zusammengedrückt und um die kleine Narbe herum stärker behaart. Die Hülsen sind breit, länglich bis linear, gerade oder gebogen, meist mit verdickten Rändern. Die schwarzen oder braunen Samen sind dick, zusammengedrückt mit einem länglichen, fleischigen Arillus.
Über 30 Arten sind in den Tropen der Alten Welt verbreitet, nur wenige in Amerika. Genutzt werden die jungen Hülsen und Samen wie bei Phaseolus-Bohnen.
Gattung Macrotyloma (Wight et Arn.) Verdc. mit 4 Arten.
Einjährige, teilweise ausdauernde Kräuter mit aufsteigendem, teilweise aufrechtem Stengel. Laubblätter mit 3 Blättchen. Die glatte Krone ist gelb, weißgelb bis grüngelb gefärbt, vereinzelt auch rötlich. Die Flügel sind schmal, das Schiffchen ist nicht gewunden. Die Staubfäden sind gleichlang und die Staubblätter zweiteilig. Der längliche bis lineare Fruchtknoten enthält 3 bis 10 Samenanlagen. Der Griffel mit der häufig schwammigen Narbe verjüngt sich nur wenig und ist schlicht, dünn behaart oder unbehaart.
Gattung Psophocarpus Neck mit 10 Arten.
Hoch windende Kräuter mit gefiederten Laubblättern, die aus drei Blättchen mit Stipellen bestehen; vielfach mit knolligen Wurzeln, die als Gemüse gegessen werden. Die häutigen Nebenblätter sind am Grunde durch einen Sporn verlängert. Die mittelgroßen Blüten stehen in Trauben in den Blattachseln. Der glockenförmige Kelch ist oben mit zwei Zipfeln verwachsen. Die Fahne ist rund und trägt am Grund nach innen gebogene Öhrchen; die eiförmigen Flügel stehen schief; das Schiffchen ist stumpf und einwärts gebogen. Die Staubfäden sind in der Mitte miteinander verwachsen. Der Fruchtknoten mit einer großen Zahl von Samenanlagen ist kurz gestielt. Die kugelige Narbe ist nach innen geneigt und ihre Oberfläche pinselförmig zottig. Die Hülsen sind vierkantig, längs der Kanten geflügelt und im Inneren zwischen den Samen mit Zellgewebe ausgefüllt. Die Samen sind länglich oval.
Im tropischen Afrika und Asien sind 5 Arten verbreitet, von denen P. tetragonolobus DC. und P. longepedunculatus Hassk. kultiviert werden.
Gattung Sphenostylis E. Mey. mit 10 Arten.
Meist windende bis niederliegende, teilweise auch aufrechte einjährige bis perennierende Kräuter, die teilweise bis über 3 m Länge erreichen. Die Wurzeln sind mit stärkereichen und eiweißhaltigen Knollen besetzt, die ebenso wie die Samen zu verschiedenen Speisen verarbeitet werden.
Die Blätter sind dreifiedrig mit langen Stielen; ihre Blättchen sind herzförmig, lanzettlich bis oval. Nebenblätter treten nicht deutlich hervor. Die zahlreichen Blüten stehen in Trauben in den Blattachseln. Sie sind gelblich bis purpurn gefärbt. Der Kelch ist leicht röhrenförmig bis glockig. Auffallend ist der extrem spitze Griffel mit zarten Haaren, der in einer abgeflachten Narbe mit scharfem Rand endet. Andere Merkmale sind nicht so deutlich verschieden. Die gesamte Pflanze ist nicht behaart.
Wahrscheinlich liegt der Ursprung der Kultur in Äthiopien. Heute sind hauptsächlich zwei Arten im tropischen Afrika verbreitet und werden dort angebaut: Sphenostylis stenocarpa (Hochst. ex Rich) Harms (Afrikanische Yambohne in Nigeria, Togo und an der Goldküste) und Sp. congensis A. Chev. (Pemo, mpempo im Kongogebiet).
Der Hauptanteil an den Leguminosensamen nimmt, wie schon dargelegt, der Embryo ein, und davon wiederum die großen, dickfleischig ausgebildeten Keimblätter, die als Speicherorgane das nur spärliche oder überhaupt nicht vorhandene Endosperm (Nährgewebe) ersetzen. Sie enthalten hauptsächlich einfache Stärkekörner, Fette und Aleuronkörner, daneben noch Dextrin, Galaktane u.a. in geringen Mengen.
Die wichtigsten Reservestoffe sind Eiweiße,
die überwiegend zu den Globulinen gehören: Albumin, Legumin,
Conglutin u.a. (s. MARQUARD: Abschnitt II und III). Bei HEGI finden sich
folgende Angaben:
Gehalt an Stickstoffsubstanz einiger Leguminosen
in %:
| Art | Rohprotein in frischen Samen | Rohprotein in Trockensubstanz | Stickstoff in der Trockensubstanz |
| Lupinus mutabilis |
|
|
|
| Lupinus luteus |
|
|
|
| Vicia faba |
|
|
|
| Pisum sativum |
|
|
|
| Lens culinaris |
|
|
|
| Cicer arietinum |
|
|
|
| Glycine hispida |
|
|
|
| Phaseolus vulgaris |
|
|
|
| Kulturart | Rohprotein | Rohfett | N-freie Extraktstoffe | Rohfaser | Asche | Wasser |
| Sojabohne
Glycine max |
33,2 | 17,5 | 30,2 | 4,4 | 4,7 | 10 |
| Erdnuß
Arachis hypogaea |
28,7 | 46,3 | 12,2 | 2,3 | 2,5 | 8 |
| Erbse
Pisum sativum |
23,4 | 1,8 | 52,5 | 5,5 | 2,8 | 14 |
| Saat-Platterbse
Lathyrus sativus |
24,7 | 2,0 | 52,2 | 5,4 | 2,7 | 13 |
| Kichererbse
Cicer arietinum |
18,6 | 5,3 | 55,4 | 4,4 | 3,3 | 13 |
| Linse
Lens culinaris |
25,5 | 1,9 | 52,2 | 3,4 | 3,0 | 14 |
| Ackerbohne
Vicia faba |
25,4 | 1,5 | 48,5 | 7,1 | 3,5 | 14 |
| Schwarze
Ackerbohne
Vicia narbonensis |
22,1 | 1,9 | 51,0 | 9,4 | 2,6 | 13 |
| Saatwicke
Vicia sativa |
26,3 | 1,7 | 49,8 | 6,0 | 3,2 | 13 |
| Ervilie
Vicia ervilia |
17,3 | 1,3 | 63,8 | 4,2 | 2,4 | 11 |
| Wicklinse
Vicia articulata |
19,3 | 2,1 | 62,5 | 3,4 | 2,7 | 10 |
| Gartenbohne
Phaseolus vulgaris |
23,5 | 1,9 | 55,4 | 3,7 | 3,5 | 12 |
| Feuerbohne
Phaseolus coccineus |
19,4 | 2,8 | 55,2 | 6,8 | 3,8 | 12 |
| Kulturart | Rohprotein | Rohfett | N-freie Extraktstoffe | Rohfaser | Asche | Wasser |
| Erdbohne
Macrotyloma geocarpum |
24,7 | 1,6 | 54,1 | 5,8 | 3,8 | 10 |
| Pferdebohne
Macrotyloma uniflorum |
25,2 | 1,0 | 56,9 | 3,9 | 3,0 | 10 |
| Flügelbohne
Psophocarpus tetragonolobus |
25,4 | 8,3 | 42,0 | 11,0 | 3,3 | 10 |
| Knollenbohne
Sphenostylis stenocarpa |
20,0 | 1,2 | 60,0 | 5,6 | 3,2 | 10 |
| Yambohne
Pachyr(r)hizus erosus |
16,3 | 0,7 | 62,7 | 6,0 | 4,3 | 10 |
| Mexikan.
Yambohne
Pachyr(r)hizus tuberosus |
20,6 | 1,4 | 60,5 | 5,7 | 2,8 | 9 |
| Rote
Spargelerbse
Lotus tetragonolobus |
25,7 | 1,7 | 50,6 | 5,7 | 5,3 | 11 |
| Bockshornklee
Trigonella foenum graecum |
24,3 | 6,0 | 48,3 | 6,3 | 4,1 | 11 |
Die Samenschale der Hülsenfrüchtler ist sehr verschieden ausgebildet, sie kann bis zu 50 % des Samengewichtes ausmachen. Sie beeinflußt im wesentlichen den Rohfasergehalt der Samen. Die Samenschale mit ihrem hohen Rohfaseranteil kann in der Tier- oder auch Humanernährung als Ballaststoff wesentliche ernährungsphysiologische Funktionen haben. Vielfach senkt sie aber den Futterwert.
Auch die Hülsen und das Stroh enthalten wertvolle Nährstoffe, die in der Tierernährung genützt werden können (Inhaltsstoffe und Nährwerte von Hülsen und Stroh siehe bei STÄHLIN). Die weiteren, den Nährwert für Mensch und Tier bestimmenden Inhaltsstoffe, besonders Rohfett und Rohfaser, variieren zwischen den Arten erheblich (s. Tab. 1).
Literatur
BECKER-DILLINGEN, J., 1929: Handbuch des Hülsenfruchterbaues und Futterbaues. Verlag Paul Parey, Berlin.
BROUWER, W. & A. STÄHLIN, 1975: Handbuch der Samenkunde. 2. Aufl., DLG-Verlag, Frankfurt/Main.
DUKE, J. A., 1981: Handbook of legumes of world economic importance. 102-106. Plenum Press, New York, London.
ENGLER, A., 1964: Syllabus der Pflanzenfamilien. 12. Aufl., Bd. II. Gebrüder Bornträger, Berlin.
ENGLER, A. & K. PRANTL, 1891: Die natürlichen Pflanzenfamilien nebst ihren Gattungen und wichtigeren Arten, insbesondere den Nutzpflanzen. III. Teil, 2. Abt. Verlag Wilhelm Engelmann, Leipzig.
FRUWIRTH, C., 1921: Handbuch des Hülsenfruchterbaues. 3. Aufl., Verlag Paul Parey, Berlin.
GEISLER, G., 1980: Körnerleguminosen. In: Pflanzenbau. 305-323. Verlag Paul Parey, Berlin.
HEGI, G., 1964: Leguminosen, Hülsenfrüchte. In: Illustr. Flora von Mitteleuropa. 2. Aufl., Bd. IV/3. Verlag Paul Parey, Berlin.
HILTNER, L., 1917: Vermehrte Futtergewinnung aus der heimischen Pflanzenwelt. Bd. I. Die Gewinnung von Futter auf dem Ackerland. Verlag Eugen Ulmer, Stuttgart.
KELLNER, O. & A. SCHEUNERT, 1952: Grundzüge der Fütterungslehre. 11. Aufl., Verlag Paul Parey, Berlin.
KIRSCH, W., K. RICHTER & K. NAUMANN, 1955: Futterwerttabellen der DLG. 2. Aufl., DLG-Verlag, Frankfurt/Main.
MANSFELD, R., 1986: Leguminosae. In: Verzeichnis landw. Kulturpflanzen. 2. Aufl., Bd. 1-4, 430-698. Springer Verlag , Berlin.
NEHRING, K., 1953: Lehrbuch der Tierernährung und Futtermittelkunde. 4. Aufl., Verlag Neumann, Radebeul und Berlin.
PURSLEGLOVE, J. W., 1968: Tropical Crops, Dicotyledons. Longmans, Green Co Ltd, London and Harlow.
RACHIE, K. O. & L. M. ROBERTS, 1974: Grain legumes of the lowland tropics. Adv. Agron. 26, 1-132.
REHM, S. & G. ESPIG, 1983: Die Kulturpflanzen der Tropen und Subtropen. 2. Aufl. Verlag Eugen Ulmer, Stuttgart.
ROYAL BOTANIC GARDENS, 1970: Kew Bulletin. Vol. 24. Her Majesty's Stationery Office, London.
SCHEIBE, A., 1953: Hülsenfruchtbau. In: ROEMER, SCHEIBE, SCHMIDT, WOERMANN: Handbuch der Landwirtschaft. 2. Aufl., Bd. II.: Pflanzenbaulehre. 248-317. Verlag Paul Parey, Berlin.
SOUCI, S. W., W. FACHMANN & H. KRAUT, 1994: In: Hülsenfrüchte und Ölsaaten. Die Zusammensetzung der Lebensmittel, Nährwert-Tabellen.4. Aufl., 743-781. CRC-Press, London, Tokio.
STÄHLIN, A., 1957: Leguminosae, Hülsenfruchtgewächse. In: Die Beurteilung der Futtermittel. Methodenbuch Bd. XII. 328-399. Verlag Neumann, Radebeul und Berlin.
TROLL, W., 1957: Praktische Einführung in die Pflanzenmorphologie. Zweiter Teil: Die blühende Pflanze. VEB Gustav Fischer Verlag, Jena.
